Раса (Бейкер, 2015)
ЗНАЧЕНИЕ «РАСЫ». Раса или подвид уже неоднократно упоминалась в главе о значении видов. Читатель должен знать, что следствием широкого распространения животного по всему ареалу его среды обитания часто могут стать изменения его внешности. Изменения особенно очевидны там, где присутствуют частичные географические барьеры, такие как цепь гор, пустыня или широкая река. Популяция по разные стороны такого барьера, однако, не полностью различна. Промежуточные формы могут быть найдены, и это часто прямое свидетельство скрещивания, хотя и не настолько масштабного, чтобы сделать две популяции неразличимыми. Если две популяции настолько различны, что можно в целом сказать, из которого региона образец был получен, то принято давать разные имена двум расам. Если каждый образец может быть с уверенностью идентифицирован как принадлежащий к одной популяции или к другой, это будет доказательством, что между ними двумя не существует миграции генов, и они поэтому могут быть рассмотрены как разные виды в генетическом смысле слова, однако различия при этом могут быть небольшими. Тот факт, что получаются промежуточные формы, определяет наличие расы. Определение любой определенной расы должно следовать от общего к частному в том смысле, что дает общее представление об отличительных характеристиках, без претензии на детальное описание любой особи.
Для практических целей представляется удобным принять произвольное решение о том, какая доля промежуточных форм допустима, если распознаются различные расы. Можно утверждать, что популяция «А» отличается от популяции «В», если «X» процентов особей, составляющих популяцию «А», может быть признано как не принадлежащие к популяции «В». Следует иметь в виду, что правильное значение, которое будет присвоено «X», не может быть обнаружено объективными средствами, тем не менее, если большая цифра (возможно — 75%) является согласованной таксономистами, можно почти не сомневаться, что существующее различие заслуживает признания в качестве подвидового или «расового». Очень часто, однако, наблюдаемые различия настолько очевидны и «X» настолько явно высока, что ни в каком
[154]
статистическом исследовании нет необходимости, чтобы убедить таксономистов, что расы следует отличать.
Для многих целей удобно рассматривать целые серии рас как кольцевые виды, но обычно принимается идея вида и система номенклатуры, базирующаяся на той, что использовал Линней, родовое и видовое имя и за ним стоит третье, подвидовое (к примеру Larus argentalus antelius). Эта трехсоставная система уже была разъяснена (глава 5) и использовалась в разных местах этой книги. Наименования рас упрощают обсуждение многих очень важных проблем, не стремясь, насколько можно видеть, никого вводить в заблуждение, утверждая, что расовые различия больше или что они более частное явление, чем есть на самом деле. Некоторые исследователи этого предмета, однако, выступают против использования трехчленной системы на том основании, что этому способу разграничения рас присуща определенная степень произвола (148). Утверждается, что один систематик может выбрать определенный набор отличительных признаков для определения двух или более рас, в то время как некоторые другие ученые той же группы могут выбрать другой набор символов и, как результат, разделить виды по-разному. Однако подозрительно, почему эта аргументация применяется более строго к расе, чем к остальным таксонам. Универсального соглашения по таксонометрическим вопросам не следует ожидать, но есть много вопросов в биологии, которые не могут быть решены до конца без помощи таксономистов. Исследователи расы готовы скорректировать свои классификации, когда для этого предоставлены достаточные основания.
Иногда утверждается, что существование промежуточных звеньев делает расы несуществующими. Но нужно отметить, что в других вопросах никто не подвергает сомнению реальность категорий, между которыми существуют промежуточные звенья. Существует, например, градация между зеленым и синим цветами, но никто не отрицает, что эти слова не должны использоваться. Таким же образом, существование юноши и человека-гермафродита не является причиной, чтобы кто-нибудь запретил использование слов «мальчик», «человек» и «женщина». Особенно неоправданно привести промежуточные звенья как противоречащее реальности рас, тогда как существование промежуточных звеньев является одним из отличительных признаков расы: если нет промежуточных звеньев, нет и самих рас. Как настаивал Кант, тем кто желает отойти от чисто академических взглядов на классификацию животных — Schulsystem *, как он ее называл, следует использовать только два таксона в описаниях животного мира: Realgattung ** и ее компонента Racen *** (см.: глава 5).
_______
* Школьная система.
** В терминологии Канта — настоящий скрещивающийся вид.
*** В терминологии Канта — узнаваемые различные формы.
[155]
Адаптация к различным средам часто приводит к эволюции рас, хотя и не существует четко определенных барьеров такого рода, расы могут быть отделены друг от друга постепенным изменением среды обитания в одной части своего ареала и более резким, из-за четко обозначенных барьеров в другой. Шри-Ланка (Цейлон) предоставляет множество таких примеров. В центре острова существует горная зона, на юго-западе низинная влажная зона, остальная часть острова находится в низинной сухой зоне. В результате многие виды млекопитающих представлены в каждой зоне отдельной расой (841, 307).
К примеру, питающиеся листьями обезьяны макаки, обычно называемые «wanderoo» (Presbytis senex), представлены в низинах сухой зоны расой (P. senex senex), отличаются большим размером и довольно темной окраской, в то время как горные — бледнее, более лохматые, имеющие коренастые формы (monticola), иногда называются «медвежьи обезьяны». Низменная влажная зона разделена широкой рекой, Кали- Гангой, которая выступает барьером в разделении рас отдельных млекопитающих. На севере можно обнаружить самого маленького wanderoo (nestor), серого, с пучком на конце хвоста, на юге реки он заменяется большим черным wanderoo (verulus), который постепенно переходит к monticola в районе пика Адама. Следует заметить, что можно обнаружить постепенное изменение признаков, когда поднимаешься в гору, и более резкие изменения при пересечении реки. Надо заметить, что различия между четырьмя расами в их типичных формах гораздо более многочисленны, чем было описано выше в нескольких словах.
Как уже было коротко упомянуто выше (глава 5), расы не обязательно в каждом случае разделены географически, иногда случается, что две расы живут на одной территории, но занимают разные среды обитания («экологические ниши») внутри них. Расы и субрасы людей, вероятно, развивались главным образом как результат частичной географической изоляции, но кое-где можно найти примеры, напоминающие «экологические расы» животных. Пигмеи (Bambutids *) Африки, живущие среди негритянского (Paleonegrid **) населения, пожалуй, попадают в эту категорию. Изоляция не полная, поскольку есть свидетельства определенного количества предков бамбутидов в палеонегридном населении.
Как указывает Т. Гексли, почти любое широко распространенное наземное животное может быть использовано в качестве обоснования существования расы, но некоторые животные подходят для этой цели лучше, чем остальные, потому что о них многое известно. Желательно использовать животное, которое было бы представлено в различных расах, легко различимых (в своих типичных формах) одна от другой морфологическими признаками, каждый из которых вызван одновре-
____
* Бамбутиды.
** Палеонегриды.
[156]

Рисунок 11. Гребенчатый тритон, Triturus cristatus cristatus, мужская (вверху) и женская особь. Фурно (372).
менным действием множества генов, имеющих кумулятивный эффект («полигены»). Также желательно должны присутствовать данные, полученные из анализа генов, каждый из которых имеет наблюдаемый эффект, и в идеале надо иметь информацию о хромосомных различиях между расами. Различия в поведении и движениях также представляют особый интерес в расовых исследованиях. По всем этим аспектам гребенчатый тритон, Triturus cristatus, приближается к идеалу. Вероятно, невозможно отыскать лучшую форму, чтобы проиллюстрировать значение расы. Будет очень хорошо, если всякий, кто профессионально пишет или говорит на данную тему, почерпнет информацию об этом обычном обитателе прудов и их окрестностей во многих частях Европы (рис. 11). Детальное рассмотрение данного вопроса приведу ниже.
Различные расы гребенчатых тритонов в основном окрашены сверху в тускло-коричневый цвет с черными пятнами и имеют желто- оранжевый живот с темными пятнами на нем. Обычно присутствуют маленькие белые пятна на тускло-коричневых боках и темное горло, тоже с белыми пятнами у многих рас. Кожа бородавчатая. Расовые различия в окраске и бородавках описаны здесь.
В отношении гребенчатого тритона для рас с коротким телом характерны, как правило, длинные конечности и, наоборот, для рас, имеющих длинное тело, характерны короткие конечности (1151). Подробная информация об этих различиях также приведена ниже.
Географическое распределение рас показано на рис. 12. Гибриды возникают там, где на одной территории расы сливаются. Возле Бухареста, например, гибриды происходят между cristarus и danubialis (1151), а в окрестностях Вены все промежуточные формы получаются между этими двумя и между ними и carnifex (183).
[157]
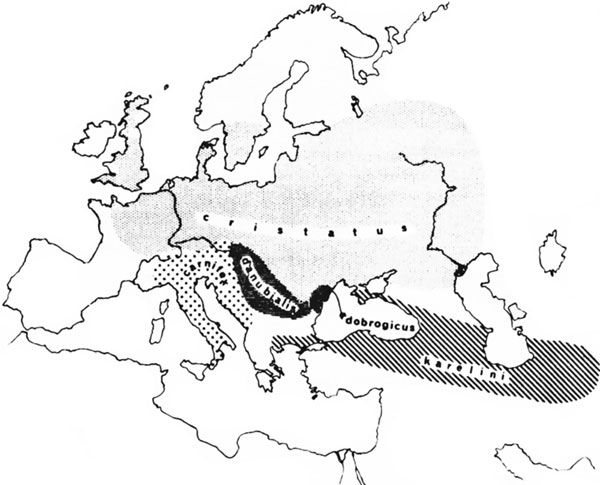
Рисунок 12. Карта Европы с указанием ареалов обитания различных рас гребенчатых тритонов Карта составлена по данным Вольтерсторфа (1151), Мертенса и Мюллера (735), Смита (982).
В неволе F1 гибриды между этими расами витальны, но F2 гибриды часто не выживают на стадии зародыша (183). Пожалуй, довольно удивительно, что гибриды смогли утвердиться в природе. Кажется, не было экспериментально установлено, что природные гибриды могут привести к бесконечно протяженной последовательности сыновних поколений.
Представители расы carnifex, которые населяют страны, прилегающие к Адриатическому морю, крупные (до 150 мм в длину) и очень коренастые, с широкой головой. Спина оказывается короткой и конечности длинные, кончики пальцев ног достигает локтя, когда тритон ставится в положение, указанное выше (см.: картинку) (1151). Верхняя поверхность имеет тенденцию к оливково-зеленому оттенку, а у самок часто встречается широкая желтая полоса посередине спины (у этого пола нет гребня у любой расы).
Благодаря исследованиям Спурвэя (1001) возможно проанализировать отдельные признаки рас генетически. Раса carnifex является рецессивной по пяти легко наблюдаемым особенностям, и это делает ее полезной в генетических исследованиях, так как доминантные гены проявляются в F1 потомстве при скрещивании. Пять генов, о которых идет речь, это:
• г, г дает достаточно ровную кожу, бородавки не очень сильно развиты,
[158]
• t, t выражается в том, что белые пятна на горле относительно малы, и эта часть кажется почти черной от цвета основы,
• s, s дает нехватку белых пятен на боках,
• I, 1 причина черных пятен на животе, отдельно расположенных друг от друга и расположенных случайным образом,
• b, b проявляется в недостатке блеска цвета берлинской лазури на коже, заметного у karelini.
Раса karelini, которая населяет часть Греции, Турции, Грузии и Ирана, может быть грубо описана с морфологической точки зрения, если сказать, что это преувеличенная форма carnifex. Он даже более массивный, голова широкая и плоская, конечности такие длинные по отношению к туловищу, что пальцы могут доставать до верха плеча. У самок никогда не бывает желтой полосы на спине.
Хотя форма тела karelini показывает, что они близкие родственники с carnifex, она несет доминантные гены пяти рецессивам последних:
• Rk, Rk производит маленькие бородавки, такие, что создается впечатление шагреневой [кожи],
• Tk, Tk дает оранжево-желтое горло, не представленное ни у одной другой расы (на нем иногда есть большие черные пятна),
• S, S дает много белых пятен на боках,
• L, L причина черных пятен на животе, сливающихся друг с другом таким образом, чтобы создать впечатление нерегулярных, темных продольных полос,
• B, B дает лазурный блеск на коже, не заметный ни у какой другой расы.
Само собой разумеется, что эта пятерка доминирующих генов, когда спаривается с пятеркой рецессивных carnifex, дает потомство F1, напоминающее себя бородавками и окраской и не очень отличающееся по форме тела, так как две расы довольно похожи в этом отношении.
По сравнению с другими расами, обычный Т. cristatus cristatus (рис. 11) Великобритании и северных частей Европы не является, с одной стороны, ни массивным, ни грацильным, с другой, и общая длина его тела (до 135 мм) является промежуточной. Пальцы выдаются вперед до основания лапы, и в этом аспекте cristatus также занимает среднюю позицию между другими расами.
• Rcr, Rcr дает крупные бородавки на коже (неясно, является ли этот ген аллеломорфным с Rk),
• Тег, Тег дает большие белые пятна на горле, которые поэтому кажутся серыми на расстоянии (неясно, является ли этот ген алломорфным с Тк),
• S, S дает белые пятна на боках, подобные тем, что у karelini,
• L, L дает неправильные, темные, продольные полосы на животе, как и у karelini,
• b, b приводит к отсутствию лазурного блеска на коже.
[159]
Долина Дуная населена другой расой, danubialis, заметно отличающейся по форме от carniflex и karelini, но демонстрирующей некоторое сходство с cristatus. Это маленький тритон (максимальная длина 125 мм), тонкий, с маленькой головой. Конечности тонкие, короткие, расставлены далеко друг от друга. Этот тритон двигается по поверхности воды по-иному, чем остальные, с сильным змеевидным изгибанием корпуса. Часто присутствует желтая полоса на спине у самок.
В своей наиболее характерной форме danubialis имеет такую генетическую конституцию:
• Rcr, Rcr дает крупные бородавки, как у типа cristatus,
• Тег, Тег дает большие белые пятна на горле, как у cristatus,
• s, s уменьшает белые пятна на боках до уровня carniflex,
• I, I предотвращает появление продольных полос на животе путем объединения черных пятен. Они округлые, резко очерченные, произвольно расположенные,
• b, b приводит к отсутствию лазурного блеска на коже.
На восточной оконечности своего ареала обитания, в районе дельты Дуная, danubialis сменяется dobrogicus, которого можно рассматривать как его преувеличенного коллегу, даже более стройного, но длиннее и с более длинными пальцами и конечностями.
Из факта, что расы скрещиваются одна с другой в природе на границах своего ареала обитания, следует, что генетическая конституция всех образцов определенных рас не одинакова.
Четыре упомянутые вначале расы все имеют по 12 пар хромосом, у dobrogicus они не были рассмотрены. Нарушения, как правило, происходят при созревании зародышевых клеток расовых гибридов. При нормальном сперматогенезе клетки в любом определенном фолликуле движутся вместе друг с другом, таким образом, они все находятся на той же самой стадии созревания в любое взятое время. Но у расовых гибридов такая синхронность нарушена и соединение отцовских и материнских наследственных хромосом часто изменено. Когда нормальное соединение отцовских и материнских наследственных хромосом предотвращается нарушениями такого рода, производятся странно выглядящие хромосомные комплексы (тривалентные, квадривалентные). Сперматогенез часто не выходит за рамки первого деления мейоза, и те, которые переживают два деления, часто вырождаются на стадии сперматида. Тем не менее, все самцы межрасовых гибридов (насколько это известно) производят некоторое количество сперматозоидов (183).
Когда Triturus cristatus скрещивается с другой расой, Т. marmoratus, сперматогенез еще более ненормальный, но разница скорее в степени, чем в виде. Расовая и специфическая разница в хромосомном поведении в сперматогенезе гибридов подтверждает степень родства, постулированную таксономистами на абсолютно различных доказательствах (632).
Если спаривание происходило между типичными представителями одинаковой расы, взятыми из мест, сильно удаленных друг от друга,
[160]

Рисунок 13. Рисунок Рюкерта хромосом типа ламповых щеток, как он их видел в развивающемся яйце морской собаки (Pristiurus) Из Рюкерта (922).
можно предположить, что нарушения могут быть найдены в сперматогенезе их потомков (1001), но этому нет подтверждений.
Когда первичные ооциты гребенчатого тритона созревают в яичниках, хромосомы становятся ненормально длинными (более чем примерно 3/4 мм) и принимают иную форму. Через их рассмотрение мы получаем представление не просто о расовых различиях, но и об их непосредственной причине, ибо «хромонема», или нить генов, отображается перед нами замечательным образом, и мы, кажется, видим в действии причины, которые приводят к различиям рас.
Эти особые хромосомы были впервые увидены в 1878 г. в развивающемся яйце аксолотля (Amblystoma tigrinum) немецким цитологом Вальтером Флеммингом и были изучены им в сотрудничестве со студентом-медиком Вибе. Их необычный вид вызвал предположение, что они могли быть артефактом, появившимся по причине влияния реагентов, использовавшихся при подготовке к изучению с помощью микроскопа. Четырьмя годами спустя Флемминг опубликовал свои результаты с некоторыми оговорками о возможности того, что вид хромосом может быть отражен неточно, не так, как при жизни (342). Между тем они видели аналогичные хромосомы у других земноводных и у некоторых неназванных рыб. Десять лет спустя гораздо более точное исследование аналогичных хромосом акулы (Pristiurus) было опубликовано еще одним немецким цитологом, Рюкертом, которому мы обязаны привычным названием, в настоящее время универсально применяемым к хромосомам Данного типа. Рюкерт писал: «Можно получить очень приблизительно гибкое представление о структуре части хромосомы, если подумать о
[161]
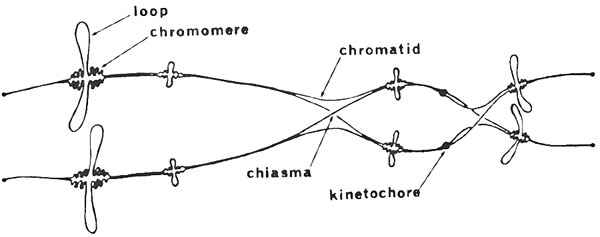
Рисунок 14. Базовая структура хромосом типа ламповых щеток. Диаграмма (приведенная не в масштабе) предназначена для представления пар этих хромосом. На каждой хромосоме отображается только четыре пары петель, но на самом деле на каждой есть сотни пар. Структура хромомеров на основе интерпретации Галла (376).
Lampen cylinderputzer *, нити которой от использования искривились и перепутались» (922). Одно из изображений Рюкерта показано на рис. 13. Название «ламповые щетки» было дано за внешнее сходство — правда, неточное, — с инструментом, использовавшимся в то время для очистки стеклянных труб масляных ламп. Он состоял из ручки, от которой во всех направлениях торчала щетина.
В настоящее время известно, что хромосомы типа ламповых щеток присутствуют в развивающихся яйцах (первичных ооцитах) большого числа животных в дополнение к земноводным и морским собакам. Они были обнаружены у определенных рептилий и птиц, у морской звезды (Echinaster), у каракатицы (Sepia) и у Anilocra, родственника мокрицы, который паразитирует на морских рыбах (180, 182). Все животные, у которых были обнаружены хромосомы типа ламповых щеток, имеют желток в яйцах. По мнению одного из ведущих авторитетов в этой области исследований, через что-то соответствующее фазе ламповых щеток, вероятно, проходят на определенном этапе созревания яйца всех животных (180).
Нашим знаниям о структуре и назначении хромосом типа ламповых щеток мы обязаны многим независимым исследователям, но особенно профессору Дж.Г. Галлу из Университета Миннесоты (374, 375, 376) и профессору Х.Г. Галлану из Университета Св. Андрея (182, 181). Из исследования фазово-контрастной и электронной микроскопии ясно, что «торчащие элементы» не такие, как у щетины в ламповых щетках, но фактически это петли продолжающейся нити ДНК (см.: рис. 14).
Каждый объект внутри ядра является парой хромосом, связанных вместе специальными соединениями (рис. 13 и 14). Каждая хромосома состоит из двух параллельных нитей и из четырех нитей (хроматиды), которые составляют целый объект — две представляют генетический
____
* Ламповая щетка.
[162]
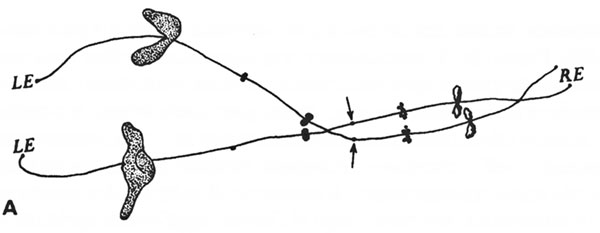
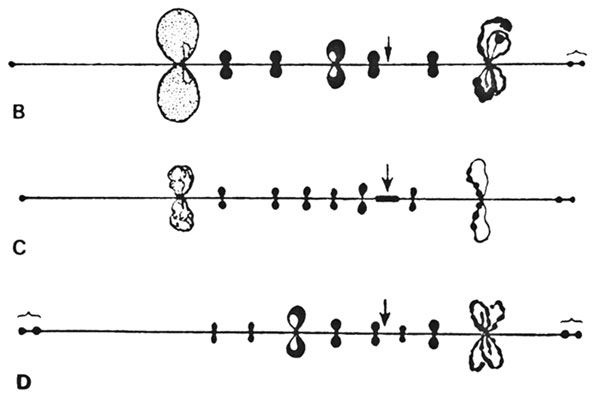
Рисунок 15. Некоторые родовые различия хромосом у гребенчатых тритонов (THturus cristatus). Диаграмма взята у Каллана и Ллойда (182).
материал матери животного, а две — происходят от отца. На диаграмме (рис. 14) показаны только 4 пары петель каждой хромосомы, фактически в некоторых хромосомах присутствуют несколько сотен.
Считается, что петли — это части хроматид, которые активны в определенный момент синтезирования генной продукции, то есть веществ, которые позволяют генам производить их эффект и последствия.
Двенадцать пар хромосом различаются по длине, и по количеству, и по расположению петли. Приблизительное постоянство в количестве и расположении петель позволяет идентифицировать каждую из пар хромосом. Они пронумерованы от I до XII, номер I самая длинная, номер XII самая короткая. Идентификация разных хромосом оказывается гораздо проще, чем могла бы быть, благодаря тому факту, что есть несколько необычных, очень толстых петель, которые настолько полезны при идентификации, что их называют «ориентиры».
Для исследователей расовых проблем существенным фактом является то, что каждая из двенадцати хромосом очень различается у разных рас Triturus cristatus. Этот предмет был изучен Калланом и Ллойдом (182), которые чрезвычайно подробно описали каждую из двенадцати хромосом типа ламповых щеток у четырех рас этого вида.
[163]
Для наших целей достаточно рассмотреть только хромосомную пару под № X *. Рис. 15, А показывает эту хромосому так, как она обычно представлена у расы carniflex, но рисунок упрощен путем пропуска всех петель, кроме ориентиров, и детали хроматиды и хиазмы не показаны. Сразу обращает на себя внимание то, что «гигантская петля» расположена в той стороне, которую можно условно назвать «левым концом» каждой хромосомы. Каллан и Ллойд (182) подготовили для каждой хромосомы то, что они назвали «рабочей картой» типичной хромосомы расы.
Рабочая карта хромосомы X carniflex показана на части «В» на рис. 15. И так же хромосома X karelini на части «С». Легко заметить основное сходство и различия, особенно намного меньший размер гигантской петли около левого конца у karelini, а также наличие утолщения оси у хромосомы этой расы. Хромосома X у cristatus показана на части «D» на том же самом рисунке. Она заметно отличается от carniflex и karelini неизменным отсутствием гигантской петли и в некоторых других аспектах, но часть ориентиров может быть распознана как соответствующая таким же у остальных рас. Гигантская петля также отсутствует в хромосоме X у danubialis.
Сравнение частей «А» и «В» на рис. 15 дает представление о количестве различий, которые могут сформироваться внутри расы.
Стрелки на рис. 15 показывают положение кинетохронов, которые задействованы при движении хромосомы при клеточном делении. Осевые перетяжки karelini фактически расположены «слева» и «справа» от кинетохронов, но не показаны отдельно.
Изучение карт полного набора хромосом гребенчатого тритона производит весьма сильное впечатление степенью различий в генетическом материале видов. Если генетическая карта будет подготовлена так, чтобы показывать все данные в двенадцати хромосомах, впечатление неизбежно будет еще более сильным.
Раса Trirurus cristatus иллюстрирует ряд важных вопросов о расовых различиях в целом. Теперь они будут рассматриваться по очереди.
1. Карты, подобные той, что представлена на рис. 12, предназначены дать представление о географическом распространении расы и не должны восприниматься слишком буквально, как указание точного ареала обитания каждой расы. Каждая территория, отмеченная по-разному, просто представляет собой ареал, внутри которого можно найти типичного представителя расы. Границы этих областей не являются резкими, как они могут казаться на карте, и они не обязательно остаются неизменными в течение длительного периода времени, происходит пересечение расовых ареалов, а промежуточные формы обычны возле границ, чем где-либо.
______
* Этот символ обозначает, что серийный номер хромосомы в аспекте длины — десять. Она не относится к «Х-хромосоме» в смысле половой хромосомы. — Прим. автора.
[164]
2. Ни один из расовых признаков не является постоянным. Помимо промежуточных гибридов есть много вариаций в размере, цвете, пропорциях частей тела, но никто не может ни спутать крайности, например dobrogicus с karelini, ни утверждать, что нашел особь любой из этих рас в дикой природе, к примеру, в Великобритании или Швеции.
3. Не все согласны с определением границ рас. Так, Волстерсторф (1151) рассматривает dobrogicus не как расу, а как субрасу danubialis, в то время как Мертенс и Мюллер (735) рассматривают их как отдельные расы. Схожим образом carniflex и karelini рассматриваются как принадлежащие к одной расе (132). Отсюда следует, что никто не может сделать уверенное заявление о числе рас или видов Triturus cristatus. То есть в случае с человеком предполагается, что вся идея с расами рушится, потому что число рас не может быть определено с уверенностью. Но факты, приведенные по отношению к расам Triturus cristatus, показывают, что это не имеющий ценности аргумент. Очевидно, что dobrogicus демонстрирует более близкое сходство с danubialis, чем с karelini. Не имеет большой важности, рассматриваем ли мы dobrogicus как субрасу danubialis или как отдельную расу, разные авторитетные мнения могут быть допущены, при настоящем уровне знаний они расходятся по этому вопросу. Серьезная ошибка, однако, будет совершена, если dobrogicus будут рассматриваться как субраса karelini или если они обе будут рассматриваться без оговорки, что принадлежат к одной расе.
Схожим образом, не имеет большого значения, будут ли carniflex и karelini рассмотрены как имеющие расовые или субрасовые различия, это будет весьма неверно разделить их и затем отнести одного из них к той же расе, что и danubialis. Опять же, балканский образец karelini более схож с carniflex, чем азиатский. Один «специалист-разделитель» может даже рассматривать балканскую и азиатскую формы как отдельные расы, а другой отнесет балканскую форму к субрасе karelini и даст ей отдельное название, но лучше быть «специалистом-объединителем» и соединить их всех под одним названием karelini. Никакой серьезной ошибки не будет при любом из этих вариантов, но, очевидно, ошибочно будет объединять балканскую форму с danubialis.
4. Расы во многих случаях различаются особенностями тела, которые могут быть переданы такими словами, как «массивность» и «грацильность». Эти термины удобны для определенных целей, но тела, указанные в вопросе, могут быть точно измерены и описаны статистическими терминами, если в этом есть необходимость. Мы упоминали массивного тритона, karelini, и его грацильного коллегу, dobrogicus. Вряд ли есть необходимость напоминать сопоставимые различия среди этнических таксонов человека. Сразу думается, например, о массивном палеонегриде из Заира и о грацильном нилотиде из Судана или о массивном альпиниде и грацильном нордиде (хотя разница между этими европидными субрасами в данном аспекте не так заметна, как между негридными субрасами). Существует, однако, огромное разнообразие этих основных
[165]
особенностей строения организма. У тритонов, например, массивность туловища связана с длинными конечностями, а грацильность — с короткими, в то время как для человека верно обратное.
Необходимо заметить, что особенности, отличающие одну расу тритонов от другой, могут быть двух видов: те, которые были проанализированы генетически, и те, которые не могут быть проанализированы таким образом. Основные морфологические особенности (форма тела, длина конечностей, к примеру) нелегко поддаются генетическому анализу. При гибридизации рас не происходит резкого разделения в аспекте таких особенностей, как эти. Всякий раз в случаях, когда разделение не наблюдается у потомства гибридов, но промежуточные формы появляются в последующих поколениях наследников, можно предположить, что изучаемые особенности обуславливаются полигенами, то есть многими генами, имеющими кумулятивный эффект. Так как различные наборы полигенов, надо полагать, обуславливают формы различных частей тела (например, различные кости и мышцы, соединенные с ними), вполне вероятно, что общее число таких генов во многих случаях больше, чем число, которое может быть проанализировано генетически. В попытке определить родство различных рас друг с другом легко понять, как неудовлетворителен тот факт, что можно описывать морфологические признаки, не имея возможности проанализировать причины, которые их создали. У некоторых животных, в частности у мушки дрозофилы, конечно, возможен генетический анализ полигенов (712), но в большинстве случаев, и прежде всего у человека, где не может быть сделано контролируемое спаривание, а размножение чрезвычайно медленное, такой анализ вряд ли возможен при существующем состоянии науки (1007, 1008) (см.: глава 9).
Идеал, к которому нужно стремиться, — это полный анализ полигенов. Пока этот идеал не будет достигнут, будет существовать ситуация, не до конца понятная исследователям (57).
5. Когда хромосомы будут исследованы на всем протяжении как хромосомы типа ламповых щеток, многочисленные генетические различия, характерные для разных рас, становятся очевидными.
6. Расы часто различаются не только формами и окрасом, но и своими привычками. Некоторые примеры приведены ниже. Triturus cristatus иллюстрируют этот тип расовых различий недостаточно хорошо, их образ жизни в общем схож, но нужно обратить внимание на очень различные плавательные действия carniflex и karelini, с одной стороны, и danubialis и dobrogicus, с другой. Это напоминает достаточно жесткие движения большинства европидов при ходьбе и гибкие движения конечностей, характерные для многих негридов. В случае с тритонами нет сомнений, что такие различия генетически обусловлены. У человека это может быть объяснено бессознательным подражанием окружению в своих действиях, но как различия могли возникнуть и распространяются на огромные районы земного шара, если генетиче-
[166]
ский элемент не принимал никакого участия? Хотя, конечно, верно, что люди любой расы могли намеренно копировать действия «своих» или избегать подражать «чужим», если они того желали.
7. В учении о расе не уделяется внимание политической карте. Пустой тратой времени будет, к примеру, собрать произвольную совокупность всех гребенчатых тритонов Румынии и описать их усредненные признаки. Конечно, можно описать усредненного «румынского» тритона. Их тела будут средней длины, ни массивными, ни гранильными, с задними конечностями, достигающими основы пальцев, он будет плавать ни в змеиной манере danubialis и dobrogicus, ни загребать, как cristatus, но совершать движения как некий промежуточный вид. Такое создание, пожалуй. может обитать в узкой зоне гибридизации, но совершенно не будет представлять великое большинство гребенчатых тритонов в Румынии, которые принадлежат к одной из трех рас, обитающих там. Любой, кто пожелает точно описать факты, вынужден будет из массы информации выделить признаки каждой расы и затем распределить их географически. Зоны гибридизации предположительно могут существовать на расовых границах, и если они могут быть точно определены, их можно нанести на карту (как, к примеру, на рис. 10). Нельзя настаивать на том, что это важная процедура, иначе существенные различия будут обязательно упущены. «Политический» метод применим только когда все образцы в определенной стране принадлежат к одной и той же расе (к примеру Т. cristatus cristatus в Швеции). Тем не менее, снова и снова данные о гетерогенных популяциях человека (особенно генетические данные) собираются и рассматриваются совершенно произвольно, только потому, что это было выгодно определенным политикам определенной нации. Информация, собранная таким образом, всегда стремится свести к минимуму и замаскировать истинные факты о расах и субрасах.
Шимпанзе, Pan satyrus (Linn.), обеспечит еще один пример расовых различий (950, 912). Читателю, возможно, будет интересно отметить характер и степень различия между расами животного, которое из всех современных видов, пожалуй, наиболее тесно связано с человеком не только морфологически, но и в отношении своих групп крови.
Недоразумения, связанные с расами шимпанзе, возникли из того факта, что голые части у юных шимпанзе всех рас телесного цвета, они начинают темнеть или становятся черными в возрасте 10—12 лет, согласно многим описаниям незрелых особей. Единственная часть, которая остается телесного цвета у старых особей, — это пенис у самцов и половые губы у самок. У юных шимпанзе всех рас в анальной области существует клок белых волос.
«Истинный» шимпанзе, Pan satyrus verus, происходит из Западной Африки, некоторые исследователи ограничивают применение имени шимпанзе только этой расой. Кожа лица у них темнеет довольно равномерно с возрастом, в результате чего нижняя часть остается бледной в течение длительного времени. Взрослый самец имеет бакенбарды и
[167]
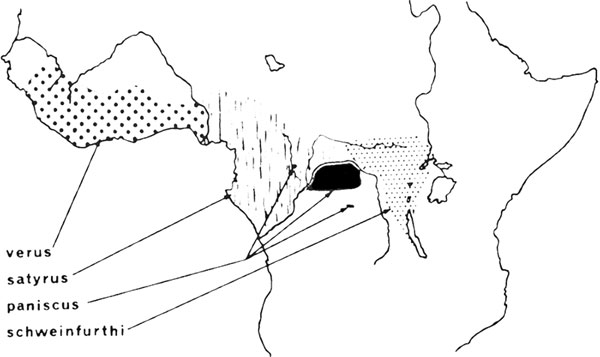
Рисунок 16. Географическое распределение (в экваториальной части Африки) рас шимпанзе Pan satyrus. Черное пятно к северу от реки Конго в ареале, занятом расой satyrus, примерно показывает то место, где образцы paniscus были захвачены, как указано в тексте. Карта нарисована по данным Родэ (912), Кулиджа (225), также Урбана и Родэ (1081).
белую бороду. Голова голая по узкой средней полосе, создает впечатление разделения на части. У черепа сильно развиты надбровные дуги, и лоб поэтому кажется сбоку заметно вогнутым. Череп высокий в теменной области и круто спускается к затылку.
К востоку от реки Нигер Pan satyrus verus сменяется лысым шимпанзе Pan satyrus satyrus, который характеризуется отсутствием волос и блестящей черной кожей на значительной части поверхности головы. Борода и бакенбарды у этой расы слабо развиты. Потемнение лица происходит нерегулярными кляксами, которые постепенно сливаются. В большинстве аспектов эта раса схожа с verus, но уши меньше, задняя часть черепа не спускается так резко до затылка, надбровные дуги сильно развиты, особенно в центральной (глабеллярной) области. Прогнатизм является более значительным, чем у verus. Эта раса была изучена в естественной среде обитания прославленным исследователем Полем дю Шайю, который назвал их Troglodytes calvus * (198, 200). Под именем Troglodytes aubryi он был впоследствии изучен анатомически очень подробно Грациоле и Аликсом, хорошо иллюстрированное описание которых было опубликовано более ста лет назад (430).
Еще одна раса шимпанзе открыта немецким исследователем Швейнфуртом на территории каннибалов Ниам-Ниам, на водоразделе
_____
* Оба Шварца и Роде (912) утверждают, что вид Du Chaillu Troglodytes koolookamba (199) фактически является Pan satyrus satyrus. Это кажется невероятным. Больше это похоже на образец verus.
[168]
рек Бахр-эль-Газаль и Убанги, и называется в его честь P. satyrus schweinfurthi. Это восточная раса, распространена до окрестностей озера Альберта в Танганьике и сильнее отличается от verus и satyrus, чем они друг от друга. Описывая в деталях анатомию этой расы почти столетие назад. Гартман отметил, что если бы он изучал его за пятнадцать лет до этого, то он выделил бы их в новый вид (476). Череп более узкий, более удлиненный, плоский сверху, профиль лица заметно прямее. Надбровные дуги не выдаются так заметно, как у других видов. Кожа лица темнеет равномерно. Мех животных очень обилен, и самец обладает хорошо развитыми бакенбардами и бородой.
Карликовый шимпанзе, Pan satyrus paniscus, был открыт много позже, чем остальные расы шимпанзе, и известен в научном мире с 1929 г. (949). Ранее предполагалось, что шимпанзе не обитают к югу от реки Конго, но эти маленькие животные довольно широко распространены в лесных регионах, ограниченных северным изгибом великой реки (947, 225) (рис. 16). В 1939 г. юный шимпанзе мужского пола был обнаружен к северу от великой реки на западе нынешней Центрально-Африканской Республики (1081, 913), то есть на территории Pan satyrus satyrus.
Pan satyrus paniscus намного меньшая обезьяна, чем verus, satyrus или schweinfurthi. Будучи взрослым, он сохраняет многие признаки, кроме размера, которые отличают молодых особей у других рас (949, 225, 1081, 913). Из них наиболее очевидные: белый анальный хохолок, маленькие уши, лоб лучше развит и голова более выпуклая в верхней части, чем у взрослых особей других рас, сзади затылок резко опускается, надбровные дуги слабо развиты, орбиты почти круговые и расположены близко друг к другу, прогнатизм мал, клыки малы, все это — юношеские признаки у других рас (хотя надо отметить, что почернение кожи лица происходит раньше у paniscus, чем у других рас).
Так как он сохраняет юношеские характеристики, будучи взрослым, Pan satyrus paniscus является примером того, что называется педоморфной * формой. Этот вопрос имеет особое значение для этой книги, поскольку он демонстрирует замечательный аналог малых народов среди человеческой популяции, например, таких, как негритиды с Малайского архипелага, пигмеи в центральной части Африки и саниды (бушмены южной части этого континента). Взрослые представители этих трех таксонов имеют примерно такое же отношение роста обычного человеческого существа с существами других человеческих таксонов, как paniscus с прочими расами шимпанзе, и все они, будучи взрослыми, демонстрируют юношеские признаки, кроме размера тела. Предмет педоморфизма обсуждается в общих чертах в главе 17, посвященной физическим характеристикам санидов.
_____
* Педоморфизм — сохранение юношеских (детских) физических черт во взрослом возрасте.
[169]
Шварц (949) считает, что paniscus ближе по некоторым своим признакам к satyrus, но Кулидж в его подробном описании относит их к schweinfurthi (225).
Расы шимпанзе отличаются друг от друга криками, которые они издают (947), но дикие антропоидные обезьяны не слишком хорошо годятся для изучения расовых различий в привычках особей. Эти животные нигде не обитают во множестве, и изучение их поведения в естественных условиях жизни всегда оказывается медленным процессом, требующим огромного терпения. Были проведены тщательные наблюдения за дикими шимпанзе, но в результате оказалось получено мало информации о различиях в привычках разных рас. Мною будет приведено несколько фактов об определенных отличиях в привычках между расами гориллы, а затем внимание будет обращено на те виды, которые более пригодны для исследований такого рода. Тщательно организованные эксперименты были проведены с дикими оленьими мышами, но, к сожалению, невозможны с ближайшими родственниками человека, шимпанзе или гориллой.
Не менее десяти видов рода Gorilla были описаны зоологами, но внимательное изучение показало, что морфологические признаки, на которых это разделение рода было основано, на самом деле очень изменчивы от особи к особи (224). Существует великое разнообразие в форме и размере сагиттального гребня и надбровных дуг и в форме затылка у этих животных. Обычно сегодня признают только один вид, Gorilla gorilla, и две расы, Gorilla gorilla gorilla и Gorilla gorilla beringei (224). Первые, так называемые «береговые» гориллы, занимают примерно ту же территорию, что и раса шимпанзе satyrus, в то время как beringei, «горные» гориллы, ограничиваются гораздо меньшей территорией, к западу от озера Эдуарда, Киву и Танганьика. У типичной горной гориллы небо длиннее, как правило, более узкий череп, руки короче, ноги длиннее, а волосяной покров гуще. Можно утверждать, что только один череп из пяти может быть идентифицирован с уверенностью как принадлежащий к той или иной форме. Те образцы горных горилл, которые были протестированы на группу крови, принадлежат к «А», а из береговых — к «Б» (189). Вероятно, пересечение некоторых признаков побудило зоологов рассматривать две формы как различающиеся виды. Поскольку их ареалы обитания не пересекаются — они отстоят примерно на 600 миль, — то не возникает вопросов о скрещивании в природе, и beringei являются носителями такой же степени родства к gorilla, как и многие «расы» животных на океанических островах к своим континентальным представителям. Поскольку генного обмена не происходит, буквальное применение определения Добжанского (глава 5) может послужить причиной того, чтобы рассматривать эти две формы как отдельные виды. Само собой разумеется, однако, что они могут скрещиваться, если бы могли встретиться в естественных условиях.
[170]
Наиболее интересные различия между двумя расами касаются их привычек. Горные гориллы, занимая возвышенности в окрестностях Митумбы, адаптированы для сопротивления холоду. Это больший объем еды и дефекация пять или шесть раз в день, они почти всегда неизбежно загаживают свое гнездо. Береговые формы едят больше фруктов, и можно предположить, что это имеет слабительный эффект, но они редко имеют дефекацию больше, чем один раз в день или гнездо делается загаженным. Из литературы по этому предмету известно, что эти привычки происходят из-за жизни в неволе. В случаях такого рода часто непросто с уверенностью провести различие между врожденными различиями, обусловленными генетической конституцией, и другими, обусловленными в основном условиями окружающей среды.
Можно привести бесчисленные примеры различий в поведении, демонстрируемые расами одного вида. Щеврица Anihus spinoletta обеспечит нас типичным примером. Водяная щеврица A. spinoletta spinoletta — птица, встречающаяся в районах болотистых лугов и карликовых деревьев в высоких горах Центральной и Южной Европы. Ее чуть меньший размер и тонкая белая полоса над глазами и по самому краю хвоста отличают ее от горной щеврицы, A. spinoletta petrosus, которая отстоит в своих привычках настолько далеко от других рас, что обитает в окрестностях скалистых морских побережий (802). Физическое сходство рас очень велико, разве что часть тела, соответствующая белой полоске у spinoletta, бледнее, чем остальное тело (но при этом не белого цвета) у petrosus.
Возникает вопрос: выбор среды обитания членов определенной расы находится ли под контролем врожденной тенденции или внешние условия обитания юной особи оставляют отпечаток в ее сознании в виде идеи «правильной» среды обитания, в которой ей необходимо проводить свою жизнь. Этот вопрос был экспериментально исследован доктором С. Уэкером из Университета Мичигана (1124, 1125). Для своих исследований он выбрал белоногую (оленью) мышь Peromyscus maniculatus. Этот вид состоит из множества рас, некоторые живут в прериях, другие — в лесных регионах. Члены рас, живущих в прериях, имеют более короткие уши и хвосты и размером несколько меньше, чем те, которые населяют леса. Раса из прерий или лугов, bairdi, и лесная раса, gracilis, обе обитали в сельской местности около Университета Мичигана, при этом каждая придерживалась специфического ареала. Задача доктора Уэкера была — обнаружить непосредственную причину выбора мышами bairdi определенной среды обитания.
Для изучения этой проблемы он выбрал участок, где росли дубовые деревья и деревья гикори, в месте, где они соприкасаются с лугом у леса, и огородил большой загон, который включал обе природные среды. Мыши были свободны внутри загона и были вольны выбрать любую среду обитания по своему предпочтению. Их перемещения автоматически фиксировались специальным устройством, которое нет необходимости здесь описывать (1124, 1125).
[171]
Дикие мыши расы bairdi, когда их помешали в загон, выбирали участок луга, предпочитая его лесу, но это мог быть результат их жизненного опыта в молодом возрасте. Поэтому дикие bairdi были перенесены в лабораторию для производства потомства. Последние не имели никакого опыта природной среды обитания, но когда их поместили в загон, они продемонстрировали очевидное предпочтение лугу. Отсюда следует, что оленьи мыши расы bairdi наследуют врожденное предпочтение их естественной среды обитания, и можно сделать вывод, что в этом отношении они отличаются от gracilis и других лесных рас.
Приведенный эксперимент, однако, не доказывает, что влияние внешних условий не имеет никакого эффекта на формирование расовых предпочтений. К счастью, возможно изучать этот эффект экспериментально. Необходимо отметить, что похожее стремление оленьей мыши прерий выбирать «правильную» окружающую среду при жизни в дикой природе поддерживается естественным отбором, потому что, когда bairdi выращивается в лабораторных условиях в течение двенадцати или более поколений, без какого-либо опыта жизни в естественной среде обитания любого рода, потомство, помещенное в экспериментальный загон, не четко демонстрирует определенные предпочтения какой-либо среды обитания, но распределяется в лесу и на лугу в случайном порядке.
Эта «лабораторная» группа мышей, у которых способность выбора среды обитания оказалась ослабленной, обеспечила материал для последующих интересных экспериментов. Некоторые из них были запущены вместе со своими матерями в закрытый загон, содержащий только луг, до того, как их глаза открылись. После примерно месяца, прожитого в этих условиях, идея луга как среды обитания была сформирована в них, и тогда, будучи перемещенными в экспериментальный загон и, таким образом, получив шанс сделать выбор между лугом или лесом, они демонстрировали предпочтение луга. Это доказывает, что в раннем возрасте внешние условия могут оказать воздействие, но эксперименты показали, что такое воздействие имеет ограниченный характер. Когда молодая особь из «лабораторной» группы была принесена в закрытый загон, содержащий только лес, перед тем, как отправиться в экспериментальный загон, содержащий луг и лес, они не демонстрировали каких-либо определенных предпочтений леса или луга. Таким образом, ослабленная «лабораторная» группа уже восстанавливала врожденную способность привычки к жизни на лугах, но не в лесу.
Этот интересный эксперимент убедительно доказывает, что генетические различия оказывают влияние на выбор среды обитания у рас оленьих мышей, хотя присутствует элемент сомнения в отношении факта, что тенденция к определенному выбору утрачивается после только 12 поколений при отсутствии естественного отбора наиболее приспособленных к выживанию. Необходимо отметить, что эксперимент был бы более наглядным, если бы он был повторен с мышами gracilis.
[172]
Расы животных отличаются тем, что может быть названо темпераментом, и есть основания предполагать, что это связано с генетикой. Было показано на опытах на крысах, что гены, влияющие на вес тела, размеры различных органов, размеры черепа и другие физические признаки, также могут влиять на послушность и покорность (178).
Особенно интересный пример расовых различий во врожденных моделях поведения обеспечивает медоносная пчела Apis mellifera *. Исследования фон Фришем «языка» этого животного хорошо известны. Благодаря его интереснейшим исследованиям, стал известным тот факт, что рабочие пчелы могут сообщить друг другу, в каком направлении и примерно на каком расстоянии находится источник еды. Нет необходимости описывать «язык», который очень понятно объяснен фон Фришем в его трудах (363, 364, 365) **.
Здесь достаточно сказать, что рабочая пчела расы A. mellifera carnica, обнаружив источник еды на некотором расстоянии от улья, возвращается домой и представляет определенный танец, который дает необходимую информацию остальным рабочим, которые вылетают из улья и находят его. Расстояние указывается скоростью, с которой танец исполняется: быстрее, значит ближе.
Существует другая раса этого вида, «итальянская» A. mellifera ligustica, которая использует, говоря метафорически — другой диалект того же самого языка. Различие в двух аспектах. Во-первых, ligustiea использует другой танец, чтобы указать направление и расстояние, когда еда расположена близко к улью. Во-вторых, для больших расстояний ее танец тот же самый, что и у carnica, но масштаб его другой: чтобы указать определенное расстояние, она танцует медленнее, чем это делает carnica. Когда особи обеих рас содержатся вместе в одном улье, они запутывают друг друга, потому что неправильно интерпретируют символы друг друга. Рабочие пчелы расы ligustiea в смешанных ульях отвечают на послание фуражиров carnica тем, что ищут пищу слишком близко к улью, и, напротив, фуражиры ligustiea посылают рабочих carnica слишком далеко в бесполезную прогулку (365).
Нет свидетельств, что юные рабочие учатся «языку» у их старших, как дети учат язык. Способность передать идею направления и расстояния врожденная, и расы врожденно различаются по этому важному компоненту поведения.
Хотя внешние условия играют такую преобладающую роль в человеческом языке, но, тем не менее, нет сомнений, что генетический элемент в нем тоже присутствует, как утверждал Эймер более чем 80 лет назад (304). Этимология и грамматика языков передаются от
_____
* Это видовое имя часто ошибочно пишется mellifica. Правила номенклатуры таковы, что должно быть соблюдено название, данное Линнеем. — Прим. автора.
** Замечательная работающая модель, объясняющая этот язык, находится в музее Оксфордского университета. — Прим. автора.
[173]
одного человека к другому в течение жизни, но фонетический элемент речи частично зависит от унаследованных признаков, которые делают простым или трудным произнести определенные звуки. Впервые это было четко установлено профессором Дарлингтоном в статье, опубликованной в генетическом журнале (247, 248), в которой он рассматривает географическое распределение по Европе людей, которые могут легко произносить различные звуки /Л-группы (как в словах this, thick и родственные звуки в языке басков), и тех, кто не может их произносить достаточно легко. Дарлингтон проследил историю человеческих миграций в связи с изменениями в распределении этой способности и пришел к заключению, что сложность в произношении этих звуков распространяется в человеческой популяции на восток. Захватчики могут принять язык своих новых подданных, сохраняя при этом свои собственные фонетические способности и неспособности. Завоеватели, таким образом, могут получить совершенно другой язык. Дарлингтон выдвигает доказательства того, что есть корреляция между низкой частотой генов группы крови «О» в популяции и трудностью в произнесении звуков «th».
Дарлингтон выражает свои выводы в национальных, а не этнических терминах, но, говоря о субрасах (с оговорками, касающимися определенных локальных групп), можно сказать, что малое количество говорящих «th» может быть найдено среди альпинидов, восточноевропейцев и арменидов, в то время как больше всего выговаривающих «th» среди нордидов и средиземноморцев.
Роль генетических факторов в развитии звуков речи была обработана в некоторых деталях Броснаханом (146).
[174]
Цитируется по изд.: Бейкер Джон Р. Раса. Взгляд белого человека на эволюцию. / Джон Р. Бейкер, перевод с английского М.Ю. Диунова. – М., 2015, с. 154-174.